
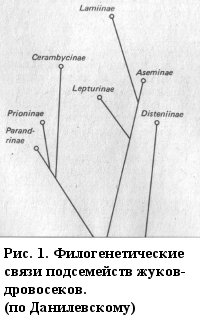
 Мировая фауна семейства делится на 9 подсемейств: Anoplodermatinae, Parandrinae, Apatophysinae, Necydalinae, Prioninae, Spondylidinae, Lepturinae, Cerambycinae, Lamiinae [Lawrence, Newton, 1995]. В России обитают представители всех подсемейств, кроме Anoplodermatinae.
Мировая фауна семейства делится на 9 подсемейств: Anoplodermatinae, Parandrinae, Apatophysinae, Necydalinae, Prioninae, Spondylidinae, Lepturinae, Cerambycinae, Lamiinae [Lawrence, Newton, 1995]. В России обитают представители всех подсемейств, кроме Anoplodermatinae.
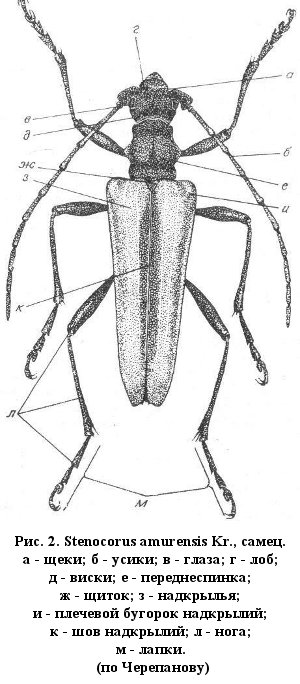 Имаго усачей отличаются более или менее вытянутым телом. Голова всегда хорошо заметна, часто довольно крупная. Её форма может быть различной, это зависит от направленности ротовых органов, степени развития и взаимного расположения усиковых бугорков, а также от размера глаз. Ротовые органы обычно направлены вперед или вперед и немного вниз. Лишь у представителей подсемейства Lamiinae они направлены вниз, иногда даже немного назад (Mesosa). Усики причленяются к верхней поверхности головы, обычно в выемках глаз, в связи с чем глаза приобретают «почковидную» форму и более или менее разделяются на верхнюю и нижнюю доли. В некоторых крайних случаях перемычка между долями глаза становится незаметной, жуки выглядят «четырехглазыми» (Tetropium, Tetrops). Глаза усачей сложные, с большим количеством отдельных фасеток. У видов, ведущих ночной или сумеречный образ жизни (Tragosoma, Prionus, Arhopalus), фасетки значительно крупнее, а число их в сложном глазу существенно меньше, чем у дневных видов.
Имаго усачей отличаются более или менее вытянутым телом. Голова всегда хорошо заметна, часто довольно крупная. Её форма может быть различной, это зависит от направленности ротовых органов, степени развития и взаимного расположения усиковых бугорков, а также от размера глаз. Ротовые органы обычно направлены вперед или вперед и немного вниз. Лишь у представителей подсемейства Lamiinae они направлены вниз, иногда даже немного назад (Mesosa). Усики причленяются к верхней поверхности головы, обычно в выемках глаз, в связи с чем глаза приобретают «почковидную» форму и более или менее разделяются на верхнюю и нижнюю доли. В некоторых крайних случаях перемычка между долями глаза становится незаметной, жуки выглядят «четырехглазыми» (Tetropium, Tetrops). Глаза усачей сложные, с большим количеством отдельных фасеток. У видов, ведущих ночной или сумеречный образ жизни (Tragosoma, Prionus, Arhopalus), фасетки значительно крупнее, а число их в сложном глазу существенно меньше, чем у дневных видов.
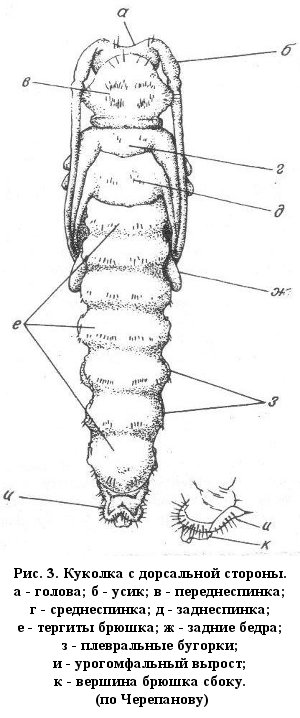 Голова представлена большой хитиновой капсулой, на переднем конце которой помещаются ротовые органы. Значительную часть головной капсулы составляют плевры, охватывающие низ, бока и часть верхней стороны капсулы. На верхней стороне капсулы, между плеврами, помещается впереди эпистом, образующий передний край капсулы. Далее сверху видны наличник, а перед ним верхняя губа. Сверху же видны челюсти, а по бокам переднего края эпистомы – короткие усики. В задней части головы имеется большая затылочная дыра (заднее отверстие головной капсулы), впереди нередко имеется и меньшее отверстие (отверстие мускулов-ретракторов). У переднего края нижней стороны головы помещается гипостом, а за ним следуют ротовые части: нижняя губа, несущая короткие щупики; по бока нижней губы – нижние челюсти с короткими парными щупиками. Нижнечелюстные щупики состоят из трех члеников, а нижнегубные всего из двух. Верхние челюсти довольно массивные, но не длинные. Глаза (простые глазки) или совсем отсутствуют, или находятся у переднего края с верхней стороны. Их бывает от 1 до 5 пар. Усики очень короткие, подогнутые книзу, состоят из трех коротких основных члеников, часто несут еще и четвертый (добавочный). Голова обычно сильно хитинизирована, а потому заметно темнее тела. Обычно голова усажена короткими, светлыми или рыжеватыми волосками. Степень втягивания головы в грудь различна у разных групп, различна и форма головной капсулы.
Голова представлена большой хитиновой капсулой, на переднем конце которой помещаются ротовые органы. Значительную часть головной капсулы составляют плевры, охватывающие низ, бока и часть верхней стороны капсулы. На верхней стороне капсулы, между плеврами, помещается впереди эпистом, образующий передний край капсулы. Далее сверху видны наличник, а перед ним верхняя губа. Сверху же видны челюсти, а по бокам переднего края эпистомы – короткие усики. В задней части головы имеется большая затылочная дыра (заднее отверстие головной капсулы), впереди нередко имеется и меньшее отверстие (отверстие мускулов-ретракторов). У переднего края нижней стороны головы помещается гипостом, а за ним следуют ротовые части: нижняя губа, несущая короткие щупики; по бока нижней губы – нижние челюсти с короткими парными щупиками. Нижнечелюстные щупики состоят из трех члеников, а нижнегубные всего из двух. Верхние челюсти довольно массивные, но не длинные. Глаза (простые глазки) или совсем отсутствуют, или находятся у переднего края с верхней стороны. Их бывает от 1 до 5 пар. Усики очень короткие, подогнутые книзу, состоят из трех коротких основных члеников, часто несут еще и четвертый (добавочный). Голова обычно сильно хитинизирована, а потому заметно темнее тела. Обычно голова усажена короткими, светлыми или рыжеватыми волосками. Степень втягивания головы в грудь различна у разных групп, различна и форма головной капсулы.
| Rhagium inquisitor (L.) | Evodinus borealis Gyllh. |
| Gnathacmaeops pratensis (Laich.) | Judolia sexmaculata (L.) |
| Strangalia attenuata (L.) | Spondylis buprestoides (L.) Gyllh. |
| Arhopalus rusticus (L.) | Asemum striatum (L.) |
| Tetropium castaneum (L.) | Molorchus minor (L.) |
| Clytus arietoides Reitt. | Monochamus urussovi (Fisch.) |
| Monochamus sutor (L.) | Monochamus saltuarius Gebl. |
| Pogonocherus fasciculatus (Deg.) | Acanthocinus griseus (F.) |
| Tragosoma depsarium (L.) | Rhagium inquisitor (L.) |
| Evodinus borealis Gyllh. | Pachyta quadrimaculata (L.) |
| Carillia virginea (L.) | Acmaeops marginatus (F.) |
| Lepturobosca virens (L.) | Judolia sexmaculata (L.) |
| Strangalia attenuata (L.) | Spondylis buprestoides (L.) |
| Arhopalus rusticus (L.) | Asemum striatum (L.) |
| Clytus arietoides Reitt. | Callidium violaceum (L.) |
| Monochamus galloprovincialis (Oliv.) | Pogonocherus fasciculatus (Deg.) |
| Acanthocinus aedilis (L.) | Acanthocinus griseus (F.) |
| Rhagium mordax (Deg.) | Stictoleptura variicornis (Dalm.) |
| Oedecnema gebleri Gangl. | Lepturalia nigripes (Deg.) |
| Macroleptura thoracica (Creutz.) | Leptura quadrifasciata L. |
| Leptura annularis F. | Stenurella melanura (L.) |
| Strangalia attenuata (L.) | Necydalis major L. |
| Xylotrechus rusticus (L.) | Cyrtoclytus capra (Germ.) |
| Rhaphuma gracilipes (Fald.) | Aegomorphus clavipes (Schr.) |
| Rhagium mordax (Deg.) | Stictoleptura variicornis (Dalm.) |
| Oedecnema gebleri Gangl. | Nivelia sanguinosa (Gyllh.) |
| Macroleptura thoracica (Creutz.) | Leptura annularis F. |
| Necydalis major L. | Aromia moschata (L.) |
| Xylotrechus rusticus (L.) | Xylotrechus pantherinus (Sav.) |
| Cyrtoclytus capra (Germ.) | Rhaphuma gracilipes (Fald.) |
| Lamia textor (L.) | Mesosa myops (Dalm.) |
| Aegomorphus clavipes (Schr.) | Saperda carcharias (L.) |
| Saperda similis (Laich.) | Saperda populnea (L.) |
| Saperda perforata (Pall.) | Oberea oculata (L.) |
| Prionus coriarius (L.) | Stenocorus meridianus (L.) |
| Nivelia sanguinosa (Gyllh.) | Macroleptura thoracica (Creutz.) |
| Rutpela maculata (Poda) | Necydalis major L. |
| Purpuricenus kaehleri (L.) | Xylotrechus arvicola (Oliv.) |
| Xylotrechus antilope (Schonh.) | Plagionotus detritus (L.) |
| Plagionotus arcuatus (L.) | Anaestetis testacea (F.) |
| Exocentrus lusitanus (L.) | Aegomorphus obscurior (Pic) |
| Pseudovadonia livida (F.) (личинки питаются разлагающимися растительными остатками, мицелием грибов и т.п.) |
| Phytoecia cylindrica (L.) |
| Phytoecia nigricornis (F.) |
| Phytoecia icterica (Shall.) |
| Agapanthia intermedia (Gangl.) |
| Усачи (Cerambycidae) | ||
| Усачи рода Monochamus | 80,3 | |
| Arhopalus rusticus (L.) | 26,9 | |
| Acanthocinus aedilis (L.) | 18,0 | |
| Rhagium inquisitor (L.) | 4,4 | |
| Короеды (Scolytidae) | ||
| Короед-стенограф (Ips sexdentatus (L.)) | 1 генерация в год | 72,0 |
| 2 генерация в год | 144,0 | |
| Малый сосновый лубоед (Tomicus minor (Hart.)) | 60,8 | |
| Короед-типограф (Ips typographus (L.)) | 1 генерация в год | 39,6 |
| 2 генерация в год | 79,2 | |
| Фиолетовый лубоед (Hylurgops palliatus (Gill.)) | 7,4 | |
| Златки (Buprestidae) | ||
| Синяя сосновая златка (Phaenops cyanea (F.)) | 46,2 | |
| Рогохвосты (Siricidae) | 39,3 |